
![]()
|
CHAPTER 5 SPECIATION UNDER THE CONDITIONS OF THE KURIL ISLANDS Charles Darwin and Alfred Wallace long ago identified the intensified rate of speciation that takes place in plants and animals on oceanic islands. This phenomenon later attracted the attention of many investigators, beginning in the middle of the 19th century, and has not lost its relevance to the present day. The evolutionary process, which comprises the adaptive transformations of populations that occur on the basis of natural selection, unfolds especially intensely on islands, under the conditions of strict isolation of island populations and a specific maritime climate. Great significance in the evolutionary process is currently attributed to microevolution, a process viewed as the aggregate of selection-directed adaptive changes within a species. The shift in the quantitative balance of forms in a polymorphous population, the formation of new morphobiological groups and ecological elements, the alteration or emergence of new populations, local ecotypes, physiological races, geographical subspecies, etc., are all considered among such changes (Zavadskiy, 1968). According to the interpretation of Timofeev-Ressovskiy et al. (1969: 61-62), "microevolution encompasses relatively small time segments; its processes are played out over limited territories and include phenomena unfolding in populations and lower taxa, concluding with speciation mechanisms." These authors advance, as "elementary evolutionary factors," the mutational process, population waves, isolation, and natural selection. All of the "elementary" factors enumerated participate in the evolutionary process, but one or several of them prove to prevail in each specific situation. Because populations are dynamic systems, when the situation changes the degree of pressure and the leading role of one or another of the evolutionary factors may change. Isolation and the specific conditions of maritime climate that lead in the aggregate to active speciation predominate on the islands. Consequently, it is necessary first of all to take note of geographical (allopatric) speciation on the islands, which is more clearly expressed here as compared with other types. Mayr (1947: 245) gives the following expanded definition for this type of speciation: "A new species is formed if any population has become geographically isolated from its parent species and has acquired, during the course of this period of isolation, characters which foster physiological isolation or guarantee it when external barriers are destroyed." Zavadskiy (1968: 315) defines geographical speciation as a "process of adaptation to new climatological and biocenotic conditions." It is unavoidably necessary when studying island floras and faunas to observe the formation of new forms and groups of organisms, both within the limits of whole archipelagos and on individual islands. The intensity of the speciation process is not identical on different islands. It depends on the antiquity of the emergence of the various islands, on the remoteness of these islands from the continents and on their origin, on the character of the relief, as well as on the attributes of the plants and animals populating the islands, factors which may result in rates of speciation that differ significantly. Speciation has by now been fairly well studied in some insects, birds, reptiles, and other plants and animals on large Pacific Ocean archipelagos (e.g., the Galápagos, Hawaiian Islands, Marquesas, Solomons, and Philippine Islands, New Guinea, etc.). As for the Kuril Archipelago, it has been little investigated from this perspective, but we have had occasion to encounter the process of speciation on Sakhalin and the Kurils when studying the entomofauna, and others (e.g., Popov, 1949; Tolmachev, 1959; Vorob'ev, 1963; Zimina and Kryshnyaya, 1968; Egorova, 1968) have had occasion to do so while studying the plants. Sakhalin, the Kuril Islands, and Japan are of continental origin. They comprised the eastern outskirts of the Asian continent in the Pliocene and possibly even later (Figure 58), and basically had a flora and fauna in common with it. Some of the plants and animals on the islands were left isolated after this territory separated from the continent. The remaking of the initial continental flora and fauna began gradually under the influence of the insular ecological conditions and the isolation that ensued. This was expressed in the emergence of subspecies and species (an especially large number of such subspecies are known among the butterflies). At the same time, some of the plants and animals were preserved unaltered under the conditions of a stable cool moist climate. The flora and fauna on the continent, where the climate became drier, were also subjected to alterations. The evolutionary process on the continent as a whole proceeded more intensely under the influence of the altered climate, and the new forms that emerged proved to be more progressive. Kuznetsov (1968b) presents data on the Kuril leaf rollers, which are interesting from this perspective: 67% of the species among the Kuril leaf rollers reveal traces of autochthonous development to one degree or another; moreover, the endemic Kuril-Japanese subspecies of the holarctic species constitute more "archaic derivatives of the Tertiary parent species than the Central Eurasian subspecies of the same species" (Kuznetsov, 1968). The Kuril subspecies of a cranefly (Tipula carinifrons violovitschi Sav.), with partially reduced wings, is more primitive by comparison with the continental subspecies of the same species (T. carinifrons gynaptera Al.), in which the wings are entirely reduced (Savchenko and Krivolutskaya, 1966). The differences and the discontinuity between the plant and animal world of the island and continental parts of the once united territory have gradually grown. This process unfolded slowly and was complicated by periodically re-occurring dry-land connections between the continent and the islands in the middle of the Quaternary period (Minato, 1966; Lebedev, 1968; Minato et al., 1968), when reciprocal migrations of plants and animals took place. This point of view is confirmed by analysis of the island entomofauna and flora, from which it can be seen that continental species or forms close to them constitute the principal nucleus of the entomofauna and flora on Sakhalin and the Kuril Islands (Table 21). Furthermore, the number of continental species is greater on Sakhalin, which had closer and later contacts with the continent, than on the Kurils. In a number of cases we find the splitting of ancient parental species into younger daughter species; a particularly large number of such examples can be cited for the insect world. For example, in ground beetles of the genus Carabus, in the section Auloncarabus, there are three very closely related species; of these, C. sichotensis Born. lives in the northern part of Sikhoté-Alin', C. diamesus Sem. et Zn. lives on Sakhalin, and C. kurilensis Lap., on the Kuril Islands and Hokkaido. They all evidently originate from the same species. The Kuril species, continuing to evolve, falls in turn into three subspecies: one restricted to the Kurils, inhabiting the islands of Iturup and Urup; and two found on Hokkaido. We are dealing here with various stages of the speciation process. There are similar examples among the leaf-beetles. There are also three closely related wingless species in the genus Chrysolina, distinguished by fairly narrow geographic ranges: C. sulcicollis Frm. is distributed in Primorskiy Kray, C. nikolskyi Jac. on Sakhalin and in Japan, and C. kabakovi L. Medv. on Kunashir and Iturup. The leaf-beetle Gonioctena chujoi L. Medv., which is very close to the Siberian G. sibirica Weise, manifests a tendency to actively speciate in different parts of its range. Three forms of it have been identified (Krivolutskaya and Medvedev, 1966; Ivliev et al., 1968): the nominative taxon on Kunashir, one subspecies G. chujoi sachalinensis L. Medv. on Sakhalin, and another subspecies G. chujoi ochotense L. Medv. on the Okhotsk littoral zone of the continent (the form that lives in Japan has not yet been described adequately, but it is not excluded that it represents a fourth). Three closely related species of long-horned grasshoppers (Metrioptera engelhardti Uv., M. japonica Bol., and M. subtropica B.-Bien.), segregated by Bei-Bienko (1954) into a separate subgenus Eobiana, also probably arose from a common ancestor. Of these, the first is known from the Russian Far East and Transbaikalia; the second from Sakhalin, Kunashir, and Hokkaido; and the third on the southern islands of Japan. A similar pattern is observed in the locusts of the genus Podismopsis, of which P. konakovi B.-Bien., an Iturup endemic, reveals kinship links with two other northern species: P. insularis Mistsh. on the Shantarskiye Islands and P. gelida Mir. in Yakutiya. Vicariants that occur in various insects and plants on the islands and
the continent attest to the same divergent transformation of initial species:
Kuznetsov (1968b) found vicariants on the continent in 15 species of Kuril
leaf-beetles.
The existence of numerous island subspecies and races, which constitute the first step in the formation of vicarious species, points to the continuing process of the divergence of the continental and island faunas. The presence of diverse ecological niches (see Chapter 4) in combination with aquatic isolation serves as a pre-condition for this on the islands. Such subspecies are encountered in nearly all groups of insects distributed on the islands; there are a particularly large number of them among the butterflies. Thus, among the diurnal butterflies (genus Rhopalocera), numbering 56 species on the Kurils, 26 (46.4%) are represented by island species. There are somewhat fewer island subspecies among other groups of butterflies, in particular among the geometrids (Geometridae); they make up 16.4%, while among the cutworm moths (Noctuidae), they make up only 6%. The activity and the antiquity of speciation on the islands can be judged by the character of the island endemism. The ancient relict forms, which are the heritage of previous faunas (usually these are clearly differentiated species), and the young new-formations (in the majority of cases, of subspecies rank or smaller intraspecies categories) must be distinguished among the island endemics. The number of palaeoendemics on the islands is relatively small. But they are represented both among the plants (Egorova, 1968) and various groups of insects (orthopterans, beetles, butterflies, dipterans, etc.). Among these are a wingless grasshopper, Diestrammena japonica Karny, which is still known only in Japan and which has vicariants on the continent; the "Lopatin" ground beetle (Captolabrus lopatini A. Mor.), found only on Sakhalin; long-horned beetles, Paraclytus excultus Bat., which belongs to an ancient genus, some representatives of which have been preserved until the present in Tibet and Western Europe, and Plectrura metallica Bat., which has been replaced on the Asian continent (the Korean Peninsula) and in North America by two other species (P. mandshurica J. Jac. and P. spinicauda Mannh.); as well as long-horned beetles which are distributed only on Kunashir and in Japan and which do not have vicariants on the continent, Doius divaricatus Bat., associated with the magnolia, Monochamus grandis Waterh., which develops on fir, and Cagosima sanguinolenta Thoms., which is associated with the alder; the bark-beetle Hyorrhynchus lewisi Blandf., which belongs to an ancient genus associated with a giant dogwood of the dogwood family (Cornaceae); and the relict leaf roller Eurydoxa advena jezoensis Mats., represented on the islands by a special subspecies, and some other species from various insect orders. These are often rare species, encountered on limited territories. Some are provisional endemics, because as the fauna and flora are more intensely studied, they are eventually found on the continent, proving once again the initial unity of the continental and island flora and fauna. For example, the bark-beetle Phellodendrophagus elegans Krivol. was described on Sakhalin, and for some time was considered an endemic of that island; we have now found it on the Southern Kuril Islands, and it was recently found in Primorskiy Kray. The ground beetle Eobruscus lutshniki Roub., known on the Southern Kurils, Southern Sakhalin, the south of Primorskiy Kray, and from the province of Hanshu in China, which undoubtedly belongs to the group of palaeoendemics, may also be regarded as a provisional endemic of the same kind. Many species of butterflies that have been regarded as Japanese have also been found on the continent in recent years. We distinguish a group of island endemics in the broad sense, based on the character of the ranges, that is, species distributed on many islands (in Japan, on the Kurils, and to some extent on Sakhalin), but which are absent on the continent. They are undoubtedly of greater age than the endemics of individual islands, and evidently arose in a period of the existence of land links between the modern islands. This is the most numerous group among the endemics: 14.8 percent among the orthopteroids, 13.4 percent among the cicads, 17.9 percent among the bugs, 12.0 percent among the ground beetles, 44.4 percent among the lamellicorns, 42.8 percent among the lycids, 34.7 percent among the click-beetles, 31.4 percent among the long-horned beetles, 17.6 percent among the leaf beetles, 11.5 percent among the bark-beetles, 16.6 percent among the bumblebees, and 7.8 percent among the flower-flies. Some of the endemic species on the Kurils are rare and are distributed only within the limits of Kunashir; others are quite common and are even numerous. Their percentage is lower on Sakhalin due to the increase there of the number of species common to the continent and the earlier disruption of the land link of Sakhalin and Hokkaido, which existed later on the Kurils. The Sakhalin-Kuril-Hokkaido endemics, which reflect the following phase of speciation, and which took place on a united territory before it broke up into individual islands, belong to a relatively younger group. This group is smaller than the proceeding in terms of the number of species, but overall comprises a characteristic complex of common or closely related forms. The following have the Sakhalin-Kuril-Hokkaido range: some beetlessubspecies of certain ground beetles (Carabus granulatus yezoensis Bat., and Cymindis subarcticus subarcticus Kano), a carrion beetle (Thanatophilus sachalinicus Kies.), a lycid (Xylobanus ogumae Mats.), click-beetles (Elater ainu Lew., Harminius singularis Lew., and Athous inornatus Lew.); many dipteranscrane flies (Tipula verrecunda Al., Nephrotoma nigricauda Al., N. minuticornis Al., Pales dorsalis sachalina Al., and Tanyptera angustistyla Al.), flower-flies (Cheilosia canicularis japonica Shir., Ch. urukavensis Shir., Ch. yesonica Mats., Neoascia longiscutata Shir., Cinxia nigriceps Shir., C. sachalinica Stack., Matsumyia nigrifacies Shir., and Takaomyia sexmaculata Mats.); a bumblebee (Bombus atrocaudatus Vogt.); and representatives of various other orders. Species with still narrower ranges, Kuril-Hokkaido (sometimes even Kunashir-Hokkaido),
Sakhalin-Hokkaido, and Sakhalin-Kuril, which reflect the connections at
different times of an initially united territory, which later broke up
gradually, belong to the same group. The Kuril-Hokkaido endemics are the
most numerous among these species; this points to the relatively recent
(evidently as recently as the late Quaternary period) interrupted link
between Hokkaido and Kunashir. There is an especially large number of such
local species and subspecies among the ground beetles, many of which are
represented by non-flying forms. For example, the island subgenus Damaster
of the genus Carabus is represented on Kunashir and Hokkaido by
C. (D.) rugipennis Motsch., while other species of
this subgenus are represented on more southerly Japanese islands. The characteristic
Kuril endemic C. (Megacarabus) aino Rost. forms two
subspecies, one of which is common to Iturup, Kunashir, and Hokkaido, while
the other lives only on Hokkaido. Such subspecies as C. opaculus opaculus
Putz., Leistus niger alecto Bat., and Agonum charillum yezoanum
Nakane are distributed only in the extreme south of the Kuril Range. Ground
beetles (Nebria jamata Motsch. and Bembidion interstrictum
Net.) that are known on the Northern Kurils and Hokkaido, but are not found
in the central and southern parts of the archipelago, are distinguished
by an interesting distribution. In addition to the ground beetles, other
insects also have the Kuril-Hokkaido range:
The Sakhalin-Kuril endemics that are unknown on Hokkaido, form a fairly
large group:
The existence of an entire group of species with such a range suggests a connection in the past between Sakhalin and the southern half of the Kuril Range that existed even after the separation of Sakhalin from the continent. Active speciation evidently took place on this territory, as a result of which endemic subspecies and species arose that were preserved after the subsidence of the central part of this island under the water only at its western and eastern margins. The endemics of the individual islands, whose final formation took place relatively recently or, as a rule, which is not yet completed, must be considered the youngest group. Among the plants and insects (Tolmachev, 1959; Vorob'ev, 1963; Egorova, 1968), the number of endemic forms on Sakhalin and the individual islands of the Kuril Range is fairly small. Morphologically they are often weakly differentiated and display great closeness to related or original species of neighboring territories (the continental part of East Asia, Japan, Kamchatka), and may sometimes coexist nearly side by side with the parent species. Such divergent speciation is encountered in some insects. For example, a grasshopper, Chorthippus kurilensis B.-Bien., is represented by well-developed subspecies on various islands: on Sakhalin (Ch.kurilensis strelkovi B.-Bien.), Kunashir (Ch. kurilensissaltator B.-Bien.), and Shikotan-Iturup (Ch. kurilensiskurilensis B.-Bien.). Its extremely close relative, Ch. fallax Zub., has an origin in common with the island species (Bei-Bienko, 1966). According to Kuznetsov (1968b), four species of island leaf rollers have island subspecies that are well differentiated by the structure of the genitalia and other characters. The Manchurian geometer Trichodezia kindermanni Brem., whose nominative form is known on Sakhalin, in China, and the Priamurye, is represented by three subspecies on the islands: Kunashir (T. kindermanni mirabilis Bryk), Hokkaido (T. kindermanni latifasciaria Prout), and Japan proper (i.e., Honshu, Shikoku, and Kyushu; T. kindermanni leechi Staudin.). Similar examples can also be cited for other widely distributed geometers that have great variability within their range:
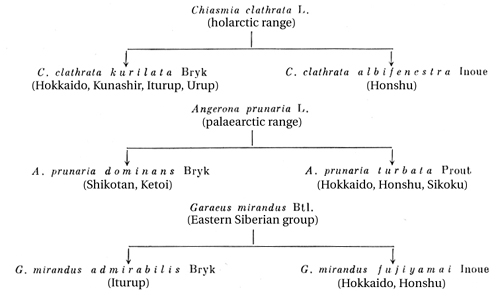
Some other species of butterflies, in particular the Maack swallowtail Papilio maacki kurilensis Mats. and the drinker Cosmotriche potatoria L., also display a tendency to form color aberrations on individual islands that are distinct from one another. Sympatric speciation is also observed on the islands along with allopatric speciation, although it appears to be substantially more difficult. For example, we have described the bark-beetle, Dryocoetes aceris Krivol., on Iturup, which is clearly a derivative of the closely related species Dryocoetes ussuriensis Egg., which, on the other hand, lives here on a different plant. Apparently, the presence on one island of two or more subspecies of the same species can be explained by sympatric speciation; this occurs in ground beetles, but also in geometers, and in other insects as well as plants. In particular, Carabus aino Rost. is represented on Hokkaido by two subspecies (C. aino chishimanus Nakane and C. aino kosugei Nakane); C. kurilensis Lap. also has two subspecies on Hokkaido (C. kurilensis daisetzuzanus Kôno and C. kurilensis rishiriensis Nakane); in the case of the geometer Anaitis perelegans kurilata Bryk, the nominative form is also encountered on Hokkaido, on a par with the Kuril-Hokkaido subspecies. Speciation that has taken place in certain local areas is highly significant in this respect; for example, the differentiation of endemic species (or local, probably ecological races) of plants close to the Moguntan mud volcano on the east coast of Sakhalin (Popov, 1949; Tolmachev, 1959). The process of speciation in the original flora and fauna remaining on the islands after their separation from one another evidently progressed more slowly, due to the stability of island climatic conditions, than on the continent, where the climate changed more rapidly in the direction of increased dryness. Therefore, the Kuril Island subspecies are sometimes found to be more archaic than those of the continental, as Kuznetsov (1968b) demonstrated in a number of leaf rollers (Tortricidae) and Savchenko and Krivolutskaya (1966) demonstrated in the craneflies (Tipulidae). But in the majority of cases, it is necessary to regard the endemics of the individual islands as the result of later, possibly even recent, speciation. Speciation has been manifested more distinctly in those organisms that reached the islands more recently, that is, in those plants and animals that had already adapted on the continent to life in different climatic conditions. It continues even at the present time. Highly interesting information is cited in the recently published studies of Zimina (Zimina and Kryshnaya, 1968) on the changes taking place in crop plants brought out from the continent and introduced into Sakhalin. The anatomical structure of organs becomes altered in peas, tomatoes, legumes, beans, and other plants after prolonged cultivation on the island: the leaves acquire a more magnocellular structure, they become thinner, the number of layers of columnar parenchyma and the thickness of their veins increases, the extent of their network decreases, the number of stomas diminishes, and their pubescence declines; enlargement of the cells takes place in the stems, and the number of layers of chlorophyllferous parenchyma and of vascular-conducting bundles increases. Consequently, the introduced plants acquire a more mesophilic structure under Sakhalin conditions. The change in external morphological structure is controlled by the internal reorganization of the plants' organism. Zimina and Berdyshev (1968a) note that a change is observed in the rhythm of development of introduced plants; the capacity for the proliferation of tissues and organs appears; the biochemical composition changes in the direction of an increased content of protein and nitrogen; and overall a delay takes place in the aging processes. Thus, introduced plants gradually acquire peculiarities that are characteristic of Sakhalin aborigines, and are eventually transformed into local Sakhalin forms. Tolmachev (1959) cites the interesting fact of the transformation under Sakhalin conditions of an annual plant, the tartary buckwheat (Fagopyrum tataricum), to a perennial semi-shrub (Fagopyrum subfructicosum) that is a morphologically and biologically new form, an endemic species, encountered nowhere other than Sakhalin. Tolmachev emphasizes that a little more than ten years was required for the transformation of the annual to the perennial, that is, only several generations. The specifics of the ecological conditionsstable elevated air humidity up to 80-85%, weakened isolation, moderate warmth, seasonal features of the climateas well as the existence of numerous local areas distinguished by a unique microclimate, and the influence of volcanic activity, are factors determining speciation on the islands besides geographical isolation. In the majority of cases we are evidently dealing with ecotypes that arise and can exist for a prolonged period of time under certain ecological conditions without becoming fixed in the genotype. When living conditions change, the characters typical of the ecotype disappear. Let us cite several examples. We found thick brakes of an onion (Allium), later described by Zimina and Kryukova (1960) as the Ichara onion, on Mount Ichara (Sakhalin's Lamonskiy mountain massif) at an altitude of 600 meters above sea level. The onion was growing in a small clearing surrounded by forest vegetation and groves of Kuril bamboo, on markedly waterlogged turf soil. The plants were remarkable for their large dimensions: the leaves reached 100 cm in length, the leaf diameter was 0.7 cm, the height of the stalks 75 cm, and the diameter of the bulb 1.5-2.0 cm. Plants brought in from the mountain were planted in fertilized meadow-sod soil in an experimental field of the Sakhalin Branch of the Academy of Sciences in Novoaleksandrovsk set. (the Susunay lowlands) and were cultivated there for several years. Starting in the first year, the dimensions of the plants began to diminish, and by 1961, when we were again able to examine them, the dimensions were halved, and they differed markedly in appearance from those growing in natural conditions. The Japanese geographical form of the spruce bark-beetle (Ips typographus f. japonicus Niiji.), which is distinguished from the type species only by the glossy, lustrous declivity of the elytra, may probably serve as another example of the ecotype. As was noted in Chapter 4, this form has been encountered in Japan, on the Kuril Islands, on Sakhalin, and in the coastal Tugurskiy Rayon of Khabarovskiy Kray, and was recently found in the alpine Ukrainian Carpathians (Pogorilyak, 1968). The lustrous declivity is also preserved in populations of the spruce bark-beetle living under conditions of elevated humidity. But even on Sakhalin, in populations living at sites remote from the sea, with a drier climate, for example in the Tym'-Poronay Valley, this character levels out and may even disappearance entirely. The gamut of transitions, from the dull elytral declivity, which is characteristic of the typical spruce bark-beetle, to the glossy lustrous declivity, is encountered here. On the continent, aside from coastal and alpine regions with a moist climate, this character is never manifested in the spruce bark-beetle. Morphological differences also appear on the Kuril Islands in the two-toothed pine wood engraver (Pityogenes bidentatus Herbst.); these are expressed in the fact that an additional pair of small sharp denticles is situated above the two hook-shaped denticles on the elytral declivity of the males. Such denticles are usually absent in the typical form on the continent, or if they are present, only on individual specimens as an exception. This character is stable in the Kuril population, since supplementary denticles are present in all males, and their size varies only somewhat (the series examined comprises several hundreds of specimens). It cannot be excluded that the formation of an ecotype is also taking place here. Deviations from the initial forms are highly diverse in different organisms, but in general they development in a direction that answer the demands of specific ecological conditions, in this case elevated atmospheric humidity, decreased and slightly fluctuating temperatures, and decreased isolation. The existence of complexes of tall herbaceous vegetation, with all the features characteristic of them, not only on the islands, but also in alpine regions and in the polar regions, is apparently explained by similar ecological conditions. Tolmachev (1944) offers an original explanation for the formation of the very large leaves of the trees growing in the Arctic in the Tertiary period. He assumes that Arctic climatic conditions at that time were characterized by relatively high summer temperatures, mild winters, an abundance of precipitation creating constant high humidity, and extreme (unbroken and at the same time thick) cloudiness. Large leaves developed in plants under such conditions as a response to the insufficient solar illumination. The large assimilation surface of the leaves made it possible to absorb a large amount of light, but large leaves could only exist under conditions of high humidity. The modern climate of Sakhalin and the Kuril Archipelago has some similarity to the climate existing in the Arctic in the Tertiary period, according to Tolmachev's concept. It is therefore entirely probable that the development of the tall herbaceous vegetation complex, which is distinguished by thick leaves (more than 1 m in diameter), was determined on the islands by the same factors, that is, by insufficient isolation at the start of vegetation, by high atmospheric humidity, and by the evenness of temperatures. Insects react to such climatic conditions in different ways. Some of them acquire melanistic coloration, and others, a glossiness of the integument; still others lose fine hairs and scales; others change the phenological development periods, sometimes forming an additional summertime diapause. Still others alter their behavior; for example, some crepuscular insects fly in the daytime hours in the presence of sunlight (see Chapter 4, Section 2). The tempo of the evolutionary process, as already noted, varies in different groups of organisms. It is generally known that progressively developing taxa are polytypic. If families, they consist of a large number of genera, while genera in their turn include a large number of species. Conversely, groups which are "degrading" in the evolutionary sense are usually represented by a small number of forms or are even monotypic. The progressiveness of various taxa is often judged on the basis of this character. The beginning stages of the establishment of species are observed in the majority of cases in the Kuril insectsthe formation of local races and subspecies, sometimes localized on individual islands. These island endemics must undoubtedly be assigned to the young island endemics. Some authors assert that in general "the endemism of the Kuril Islands in terms of its taxonomic significance does not rise above subspecies categories" (Klumov, 1962: 38). However, that is not always the case. Among the insects studied on the islands, some groups display the capacity for intensive speciation. A small family of beetles characterized by taxa that fly quite poorly, the Lycidae, may serve as an example. We have already touched upon this question when we reviewed the family Lycidae in Chapter 3. Let us briefly recall the facts presented there. The family Lycidae, which apparently originated in the tropical regions of East Asia, is in general distinguished by active speciation, and forms a large number of morphologically similar species, both on the continent (30 species in China) and on the islands (22 in Japan). This family is represented on the Southern Kuril Islands by 21 species, of which eight are Kuril endemics and six are endemics of several islands (Sakhalin, Japan, and the Kurils). Furthermore, some of the Kuril species display kinship connections with other island species, and evidently are a derivative of them, since as a rule the ranges of these closely related forms overlap. A process of recent speciation that has reached a concluding stage is undoubtedly taking place here, that is, of differentiation into individual species. Analogous phenomena may be found in other groups of insects as well, but which are insufficiently studied in this respect. Summarizing all the above, it can be concluded that the limited number of endemic forms on particular islands, their low level of differentiation, and the actively proceeding processes of recent speciation in various groups of organisms attest to the relatively recent occurrence of insular isolation. This is also confirmed by the latest geological data (Minato, 1966; Lebedev, 1968). It is also germane to stress that by contrast with oceanic islands, the processes of speciation on the continental islands of East Asia after their separation from the continent apparently proceeded more slowly than on the continent, due to the prolonged maintenance of climatic conditions that are close to the initial conditions. The study of the speciation of plants, insects, and vertebrates on Sakhalin, the Kuril Islands, and evidently in Japan remains far from adequate. We are able to make judgments regarding this important process only on the basis of isolated observations and on the basis of those small experimental investigations of botanists that have been mentioned. Most of the reasoning that has been advanced by various scientists on this score is purely speculative. A wide range of special investigations, including a large group of scientists of various specialties setting up experiments, is required to resolve this extremely interesting problem. | |||||||||||||||||||||
![]()